What is a Cell?
Cells are the structural and functional unit of all living organisms. Some organisms, like bacteria, are unicellular-consisting of a single cell. Other organisms, such as humans, are multicellular, or have many cells--an estimated 100,000,000,000,000 cells! Each cell is an amazing world onto itself: it can take in nutrients, convert these nutrients into energy, carry out specialized functions, and reproduce as necessary. Even more amazing is that each cell stores its own set of instructions for carrying out each of these activities.
Cell Organization
Before we can discuss the various components of a cell, it is important to know what organism the cell comes from. There are two general categories of cells: prokaryotes and eukaryotes.
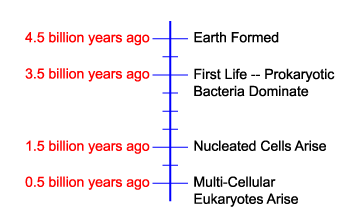
History of Life On Earth
Prokaryotic Organisms
It appears that life arose on earth about 4 billion years ago. The simplest of cells, and the first types of cells to evolve, were prokaryotic cells--organisms that lack a nuclear membrane, or the membrane that surrounds the nucleus of a cell. Bacteria are the best known and most studied form of prokaryotic organisms, although the recent discovery of a second group of prokaryotes, called archaea, has provided evidence of a third cellular domain of life and new insights into the origin of life itself.
Prokaryotes are unicellular organisms that do not develop or differentiate into multicellular forms. Some bacteria grow in filaments, or masses of cells, but each cell in the colony is identical and capable of independent existence. The cells may be adjacent to one another because they did not separate after cell division or because they remained enclosed in a common sheath or slime secreted by the cells. Typically though, there is no continuity or communication between the cells. Prokaryotes are capable of inhabiting almost every place on the earth, from the deep ocean, to the edges of hot springs, to just about every surface of our bodies.
Prokaryotes are distinguished from eukaryotes on the basis of nuclear organization, specifically their lack of a nuclear membrane. Prokaryotes also lack any of the intracellular organelles and structures that are characteristic of eukaryotic cells. Most of the functions of organelles, such as mitochondria, chloroplasts and the Golgi apparatus, are taken over by the prokaryotic plasma membrane. Prokaryotic cells have three architectural regions: appendages called flagella and pili--proteins attached to the cell surface; a cell envelope consisting of a capsule, a cell wall and a plasma membrane; and a cytoplasmic region that contains the cell genome (DNA) and ribosomes and various sorts of inclusions.
Eukaryotic Organisms
Eukaryotes include fungi, animals, and plants as well as some unicellular organisms. Eukaryotic cells are about 10 times the size of a Prokaryote and can be as much as 1000 times greater in volume. The major and extremely significant difference between prokaryotes and eukaryotes is that eukaryotic cells contain membrane-bounded compartments in which specific metabolic activities take place. Most important among these is the presence of a nucleus, a membrane-delineated compartment that houses the eukaryotic cell’s DNA. It is this nucleus that gives the eukaryote--literally, true nucleus--its name.
Eukaryotic organisms also have other specialized structures, called organelles, which are small structures within cells that perform dedicated functions. As the name implies, you can think of organelles as small organs. There are a dozen different types of organelles commonly found in eukaryotic cells. In this primer we will focus our attentions on only a handful of organelles and will examine these organelles with an eye to their role at a molecular level in the cell.
The origin of the eukaryotic cell was a milestone in the evolution of life. Although eukaryotes use the same genetic code and metabolic processes as prokaryotes, their higher level of organizational complexity has permitted the development of truly multicellular organisms. Without eukaryotes, the world would lack mammals, birds, fish, invertebrates, mushrooms, plants, and complex single-celled organisms.

Figure 2. Eukaryotes and Prokaryotes
--------------------------------------------------------------------------------
This figure illustrates a typical human cell (eukaryotic cell) and
a typical bacterium (prokaryotic cell). The cartoon on the left highlights
the internal structures of eukaryotic cells, including the nucleus (light
blue), the nucleolus (intermediate blue), mitochondria (orange), and ribosomes
(dark blue). The drawing on the right demonstrates how bacterial DNA is
housed in a structure called the nucleoid (very light blue), as well as
other structures normally found in a prokaryotic cell, including the cell
membrane (black), the cell wall (intermediate blue), the capsule (orange),
ribosomes (dark blue), and a flagellum (also black).
--------------------------------------------------------------------------------
Cell Structures: The Basics
The Plasma Membrane--A Cell's Protective Coat
The outer lining of a eukaryotic cell is called the plasma membrane. This membrane serves to separate and protect a cell from its surronding environment and is made mostly from a double layer of proteins and lipids--fat-like molecules. Embedded within this membrane are a variety of other molecules that act as channels and pumps, moving different molecules into and out of the cell. A form of plasma membrane is also found in prokaryotes, but in this organism it is usually referred to as the cell membrane.
The Cytoskeleton--A Cell's Scaffold
The cytoskeleton is an important, complex, and dynamic cell component. It acts to organize and maintain the cell's shape; anchors organelles in place; helps during endocytosis--the uptake of external materials by a cell; and moves parts of the cell in processes of growth and motility. There are a great number of proteins associated with the cytoskeleton, each controlling a cell’s structure by directing, bundling and aligning filaments.
The Cytoplasm--A Cell's Inner Space
Inside the cell there is a large fluid-filled space called the cytoplasm, sometimes called the cytosol. In prokaryotes, this space is relatively free of compartments. In eukaryotes, the cytosol is the "soup" within which all the cell's organelles reside. It is also the home of the cytoskeleton. The cytosol contains dissolved nutrients, helps break down waste products, and moves material around the cell through a process called cytoplasmic streaming. The nucleus often flows with the cytoplasm changing its shape as it moves. The cytoplasm also contains many salts and is an excellent conductor of electricity, creating the perfect environment for the mechanics of the cell. The function of the cytoplasm, and the organelles which reside in it, are critical for a cell's survival.
Genetic Material
Two different kinds of genetic material exist; deoxyribonucleic acid (DNA) and ribonucleic acid (RNA). Most organisms are made of DNA, but a few viruses have RNA as their genetic material. The biological information contained in an organism is encoded in its DNA or RNA sequence.
Prokaryotic genetic material is organized in a simple circular structure that rests in the cytoplasm. Eukaryotic genetic material is more complex and is divided into discrete units called genes. Human genetic material is made up of two distinct components: the nuclear genome and the mitochondrial genome. The nuclear genome is divided into 24 linear DNA molecules, each contained in a different chromosome. The mitochondrial genome is a circular DNA molecule separate from the nuclear DNA. Although the mitochondrial genome is very small, it codes for some very important proteins.
Organelles
The human body contains many different organs, such as the heart, lung, and kidney--with each organ performing a different function. Cells also have a set of "little organs," called "organelles," that are adapted and/or specialized for carrying out one or more vital functions. Organelles are found only in eukaryotes and are always surrounded by a protective membrane. It is important to know some basic facts about the following organelles.
The Nucleus--A Cell's Center
The nucleus is the most conspicuous organelle found in a eukaryotic cell. It houses the cell's chromosomes and is the place where almost all DNA replication and RNA synthesis occurs. The nucleus is spheroid in shape and separated from the cytoplasm by a membrane called the nuclear envelope. The nuclear envelope isolates and protects a cell's DNA from various molecules that could accidentally damage its structure or interfere with its processing. During processing, DNA is transcribed, or synthesized into a special RNA, called mRNA. This mRNA is then transported out of the nucleus where it is translated into a specific protein molecule. In prokaryotes, DNA processing takes place in the cytoplasm.
The Ribosome--The Protein Production Machine
Ribosomes are found in both prokaryotes and eukaryotes. The ribosome is a large complex comprised of many molecules, including RNAs and proteins, and is responsible for processing the genetic instructions carried by a mRNA. The process of converting a mRNA's genetic code into the exact sequence of amino acids that make up a protein is called translation. Protein synthesis is extremely important to all cells, and so a large number of ribosomes--sometimes hundreds or even thousands--can be found throughout a cell.
Ribosomes float freely in the cytoplasm, or sometimes bound to another organelle called the endoplasmic reticulum. Ribosomes do not have a disassemble into subunits--a large and a small--when not actively synthesizing proteins.
Mitochondria and Chloroplasts--The Power Generators
Mitochondria are self-replicating organelles that occur in varying numbers, shapes, and sizes in the cytoplasm of all eukaryotic cells. As mentioned earlier, mitochondria contain their own genome that is separate and distinct from the nuclear genome of a cell. Mitochondria have two functionally distinct membrane systems separated by a space: the outer membrane, which surrounds the whole organelle, and the inner membrane that is thrown into folds or shelves that project inward. These inward folds are called "cristae." The number and shape of cristae in mitochondria differs depending on the tissue and organism in which they are found and serve to increase the surface area of the membrane.
Mitochondria play a critical role in generating energy in the eukaryotic cell and this process involves a number of complex pathways. Lets break down each of these steps so you can better understand how food and nutrients are turned into energy packets and water. First, some of the best energy supplying foods we eat contain complex sugars. These complex sugars can be broken down into a less chemically complex sugar molecule called glucose. Glucose can then enter the cell through special molecules found in the membrane, called "glucose transporters". Once inside the cell, glucose is broken down to make adenosine triphosphate, or ATP--a form of energy--via two different pathways.
The first pathway, glycolysis, requires no oxygen and is referred to as anaerobic metabolism. Glycolysis occurs in the cytoplasm outside the mitochondria. During glycolysis, glucose is broken down into a molecule called pyruvate. Each reaction is designed to produce some hydrogen ions that can then be used to make energy packets (ATP). However, only four ATP molecules can be made fromone molecule of glucose run through this pathway. In prokaryotes, glycolysis is the only method used for converting energy.
The second pathway, called the Kreb's cycle, or the citric acid cycle, occurs inside the mitochondria and is capable of generating enough ATP to run all the cell functions. Once again, the cycle begins with a glucose molecule, which during the process of glycolysis, was stripped of some of its' hydrogen atoms, transforming the glucose into two molecules of pyruvic acid. Next, pyruvic acid is altered by the removal of a carbon and two oxygens, which go on to form carbon dioxide. When the carbon dioxide is removed, energy is given off, and a molecule called NAD+ is converted into the higher energy form NADH. Another molecule, coenzyme A, then attaches to the remaining acetyl unit, forming acetyl CoA.
Acetyl CoA enters the Kreb's cycle by joining to a four-carbon molecule called oxaloacetate. Once the two molecules are joined, they make a six-carbon molecule called citric acid. Citric acid is then broken down and modified in a stepwise fashion. As this happens, hydrogen ions and carbon molecules are released. The carbon molecules are used to make more carbon dioxide. The hydrogen ions are picked up by NAD and another molecule called flavin-adenine dinucleotide, or FAD. Eventually, the process produces the four-carbon oxaloacetate again, ending up where it started off. All in all, the Kreb's cycle is capable of generating between 24 and 28 ATP molecules out of one molecule of glucose converted to pyruvate. So, it is easy to see how much more energy we can get out of a molecule of glucose if our mitochondria are working properly and if we have oxygen.
Chloroplasts are similar to mitochondria, but are found only in plants. Both organelles are surronded by a double membrane with an intermembrane space; both have their own DNA and are involved in energy metabolism; and both have reticulations, or many foldings, filling their inner space. Chloroplasts convert light energy from the sun into ATP through a process called photosynthesis.
The Endoplasmic Reticulum and the Golgi Apparatus--Macromolecule Managers
The endoplasmic reticulum (ER) is the transport network for molecules targeted for certain modifications and specific destinations, as compared to molecules that will float freely in the cytoplasm. The ER has two forms: the rough ER and the smooth ER. The rough ER is labeled as such because it has ribosomes adhering to its outer surface, while the smooth ER does not. Translation of the mRNA for those proteins that will either stay in the ER or be exported-moved out-of the cell occurs at the ribosomes attached to the rough ER. The smooth ER serves as the recipient for those proteins synthesized in the rough ER. Proteins to be exported are passed to the Golgi apparatus--sometimes called a Golgi body or Golgi complex--for further processing, packaging and transport to a variety of other cellular locations.
Lysosomes and Peroxisomes--The Cellular Digestive System
Lysosomes and peroxisomes are often referred to as the garbage disposal system of a cell. Both organelles are somewhat spherical and bound by a single membrane and rich in digestive enzymes-- naturally occurring proteins that speed up biochemical processes. For example, lysosomes can contain more than three-dozen enzymes for degrading proteins, nucleic acids, and certain sugars called polysaccharides. All these enzymes work best at a low pH, reducing the risk that these enzymes will digest their own cell should they somehow escape from the lysosome. Here we can see the importance behind compartmentalization of the eukaryotic cell. The cell could not house such destructive enzymes if they were not contained in a membrane-bound system.
One function of a lysosome is to digest foreign bacteria that invade a cell. Other functions include helping to recycle receptor proteins and other membrane components and degrading worn out organelles such as mitochondria. Lysosomes can even help repair damage to the plasma membrane by serving as a membrane patch, sealing the wound.
Peroxisomes function to rid the body of toxic substances, like hydrogen peroxide, or other metabolites and contain enzymes concerned with oxygen utilization. High numbers of peroxisomes can be found in the liver where toxic byproducts are known to accumulate. All of the enzymes found in a peroxisome are imported from the cytosol. Each enzyme transferred to a peroxisime has a special sequence at one end of the protein, called a PTS or peroxisomal targeting signal, that allows the protein to be taken into that organelle where they then function to rid the cell of toxic substances.
Peroxisomes often resemble a lysosome. However, peroxisomes are self replicating whereas lysosomes are formed in the Golgi complex. Peroxisomes also have membrane proteins that are critical for various functions, such as for importing proteins into their interiors and to proliferate and segregate into daughter cells.
Where Do Viruses Fit?
Viruses are not classified as cells and therefore are neither unicellular nor multicellular organisms. Most people do not even classify viruses as "living" as they lack a metabolic system and are dependent on the host cells that they infect to reproduce. Viruses have genomes that consist of either DNA or RNA, and there are examples of viruses that are either double-stranded or single-stranded. Importantly, their genomes code not only for the proteins needed to package its genetic material but for those proteins needed by the virus to reproduce during its infective cycle.
Making New Cells and Cell Types
For most unicellular organisms, reproduction is a simple matter of cell duplication, also known as replication. But for multicellular organisms, cell replication and reproduction are two separate processes. Multicellular organisms replace damaged or worn out cells through a replication process called mitosis--the division of a eukaryotic cell nucleus to produce two identical daughter nuclei. To reproduce, eukaryotes must first create special cells called gametes--eggs and sperm--that then fuse to form the beginning of a new organism. Gametes are but one of the many unique cell types that multicellular organisms require in order to function as a complete organism.
Making New Cells
Most unicellular organisms create their next generation by replicating all of their parts and then splitting into two cells - a type of asexual reproduction called binary fission. This process spawns not just two new cells, but also two new organisms. Multicellullar organisms replicate new cells in much the same way. For example, we produce new skin cells and liver cells by replicating the DNA found in that cell through mitosis. Yet, producing a whole new organism requires sexual reproduction, at least for most multicellular organisms. In the first step, specialized cells called gametes--eggs and sperm--are created through a process called meiosis. Meiosis serves to reduce the chromosome number for that particular organism by half. In the second step, the sperm and egg join to make a single cell, which restores the chromosome number. This joined cell then divides and differentiates into different cell types that eventually form an entire functioning organism.
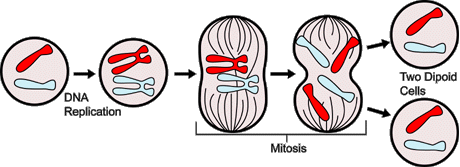
Figure 3. Overview of the Major Events In Mitosis
--------------------------------------------------------------------------------
Mitosis is the process by which the diploid nucleus-having two sets
of homologous chromosomes-of a somatic cell divides to produce two daughter
nuclei, both of which are still diploid. The left hand side of the cartoon
demonstrates how the parent cell duplicates its chromosomes (one red and
one blue), providing the daughter cells with a complete copy of genetic
information. Next, the chromosomes align at the equatorial plate and the
centromeres divide. The sister chromatids then separate, becoming two
diploid daughter cells, each with one red and one blue chromosome.
--------------------------------------------------------------------------------
Mitosis
Every time a cell divides, it must ensure that its DNA is shared between the two daughter cells. Mitosis is the process of "divvying up" the genome between the daughter cells. In order to easier describe this process, let's imagine a cell with only one chromosome. Before a cell enters mitosis, we say the cell is in interphase, the state of a eukaryotic cell when not undergoing division. Every time a cell divides, it must first replicate all its DNA. Since chromosomes are simply DNA wrapped around protein, the cell replicates its chromosomes too. These two chromosomes, positioned side by side, are called sister chromatids, and are identical copies of one another. Before this cell can divide, it must separate these sister chromatids from one another. To do this, the chromosomes have to condense. This stage of mitosis is called prophase. Next, the nuclear envelope breaks down and a large protein network, called the spindle, attaches to each sister chromatid. The chromosomes are now aligned perpendicular to the spindle in a process called metaphase. Next, molecular motors pull the chromosomes away from the metaphase plate to the spindle poles of the cell. This is called anaphase. Once this process is completed, the cells divide, the nuclear envelope reforms, and the chromosomes relax and decondense during telophase. The cell can now replicate its DNA again during interphase and go through mitosis once more.
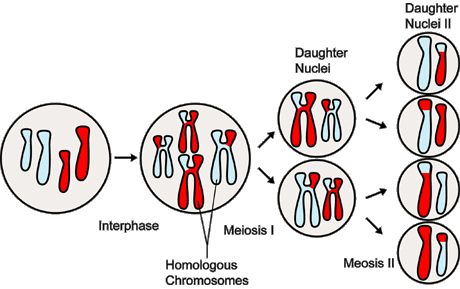
Figure 4. Overview of the Major Events In Meiosis
--------------------------------------------------------------------------------
Meiosis, a type of nuclear division, occurs only in reproductive cells
and results in a diploid cell--having two sets of chromosomes--giving
rise to four haploid cells--having a single set of chromosomes. Each haploid
cell can subsequently fuse with a gamete of the opposite sex during sexual
reproduction. In this illustration, two pairs of homologous chromosomes
enter Meiosis I, which results initially in two daughter nuclei, each
with two copies of each chromosome. These two cells then enter Meiosis
II, producing four daughter nuclei, each with a single copy of each chromosome.
--------------------------------------------------------------------------------
Meiosis
Meiosis is a specialized type of cell division that occurs during the formation of gametes. Although meiosis may seem much more complicated than mitosis, it is really just two cell divisions in sequence. Each of these sequences maintains strong similarities to mitosis.
Meiosis I refers to the first of the two divisions and is often called the reduction division. This is because it is here that the chromosome compliment is reduced from diploid--two copies-to haploid-one copy. Interphase in meiosis is identical to interphase in mitosis. At this stage, there is no way to determine what type of division the cell will undergo when it divides. Meiotic division will only occur in cells associated with male or female sex organs. Prophase I is virtually identical to prophase in mitosis, involving the appearance of the chromosomes, the development of the spindle apparatus, and the breakdown of the nuclear membrane. Metaphase I is where the critical difference occurs between meiosis and mitosis. In mitosis, all the chromosomes line up on the metaphase plate in no particular order. In Metaphase I, the chromosome pairs are aligned on either side of the metaphase plate. It is during this alignment that the chromatid arms may overlap and temporarily fuse, resulting in what is called crossovers. During Anaphase I, the spindle fibers contract, pulling the homologous pairs away from each other and toward each pole of the cell. In Telophase I, a cleavage furrow typically forms, followed by cytokinesis--the changes that occur in the cytoplasm of a cell during nuclear division, but the nuclear membrane is usually not reformed and the chromosomes do not disappear. At the end of Telophase I, each daughter cell has a single set of chromosomes, half the total number in the original cell, that is, while the original cell was diploid, the daughter cells are now haploid.
Meiosis II is quite simply a mitotic division of each of the haploid cells produced in Meiosis I. There is no Interphase between Meiosis I and Meiosis II and the latter begins with Prophase II. At this stage, a new set of spindle fibers form and the chromosomes begin to move toward the equator of the cell. During Metaphase II, all the chromosomes in the two cells align with the metaphase plate. In Anaphase II, the centromeres split and the spindle fibers shorten, drawing the chromosomes toward each pole of the cell. In Telophase II, a cleavage furrow develops, followed by cytokinesis and the formation of the nuclear membrane. The chromosomes begin to fadeand is replaced by the granular chromatin characteristic of interphase. When Meiosis II is complete, there will be a total of four daughter cells, each with half the total number of chromosomes as the original cell. In the case of male structures, all four cells will eventually develop into sperm cells. In the case of the female life cycles in higher organisms, three of the cells will typically abort, leaving a single cell to develop into an egg cell which is usually much larger than a sperm cell.
Recombination--The Physical Exchange of DNA
All organisms suffer a certain number of small mutations, or random changes in a DNA sequence, during the process of DNA replication. These are called spontaneous mutations and occur at a rate characteristic for that organism. Genetic recombination refers more to a large-scale rearrangement of a DNA molecule. This process involves pairing between complementary strands of two parental duplex, or double-stranded DNAs, and results from a physical exchange of chromosome material.
The position at which a gene is located on a chromosome is called a locus. In a given individual, one might find two different versions of this gene at a particular locus. These alternate gene forms are called "alleles." During Meiosis I, when the chromosomes line up along the metaphase plate, the two strands of a chromosome pair may physically cross over one another. This may cause the strands to break apart at the crossover point and reconnect to the other chromosome, resulting in the exchange of part of the chromosome.
Recombination results in a new arrangement of maternal and paternal alleles on the same chromosome. Although the same genes appear in the same order, the alleles are different. This process explains why offspring from the same parents can look so different. In this way, it is theoretically possible to have any combination of parental alleles in an offspring, and the fact that two alleles appear together in one offspring does not have any influence on the statistical probability that another offspring will have the same combination. This theory of "independent assortment" of alleles is fundamental to genetic inheritance. However, having said that, there is an exception that requires further discussion.
First, the frequency of recombination is actually not the same for all gene combinations. This is because recombination is greatly influenced by the proximity of one gene to another. If two genes are located close together on a chromosome, the likelihood that a recombination event will separate these two genes is less than if they were farther apart. Linkage describes the tendency of genes to be inherited together as a result of their location on the same chromosome. Linkage disequilibrium describes a situation in which some combinations of genes or genetic markers occur more or less frequently in a population than would be expected from their distance apart. Scientists apply this concept when searching for a gene that may cause a particular disease. They do this by comparing the occurrence of a specific DNA sequence with the appearance of a disease. When they find a high correlation between the two, they know they are getting closer to finding the appropriate gene sequence.
Binary Fission--How Bacteria Reproduce
Bacteria reproduce through a fairly simple process called binary fission, or the reproduction of a living cell by division into two equal, or near-equal parts. As just noted, this type of asexual reproduction theoretically results in two identical cells. However, bacterial DNA has a relatively high mutation rate. This rapid rate of genetic change is what makes bacteria capable of developing resistance to antibiotics and helps them exploit invasion into a wide range of environments.
Like more complex organisms, bacteria also have mechanisms for exchanging genetic material. Though not equivalent to sexual reproduction, the end result is that a bacterium contains a combination of traits from two different "parental" cells. Three different modes of exchange have thus far been identified in bacteria.
Conjunction involves the direct joining of two bacteria, which allows their circular DNAs to undergo recombination. Bacteria can also undergo transformation by absorbing remnants of DNA from dead bacteria and integrating these fragments into their own DNA. Lastly, bacteria can exchange genetic material through a process called transduction, in which genes are transported into and out of the cell by bacterial viruses--called bacteriophages--or by plasmids--an autonomous self-replicating extra-chromosmal circular DNA.
Viral Reproduction
Because viruses are acellular and do not use ATP, they must utilize the machinery and metabolism of a host cell to reproduce. For this reason, viruses are called obligate intracellular parasites. Before a virus has entered a host cell, it is called a virion--a package of viral genetic material. Virions--infectious viral particles--can be passed from host to host either through direct contact or through a vector, or carrier. Inside the organism, the virus can enter a cell in various ways. Bacteriophages--bacterial viruses--attach to the cell wall surface in specific places. Once attached, enzymes make a small hole in the cell wall and the virus injects its DNA into the cell. Other viruses (like HIV) enter the host via endocytosis, the process whereby cells take in material from the external environment. After entering the cell, the virus's genetic material begins the destructive process of taking over the cell and forcing it to produce new viruses.
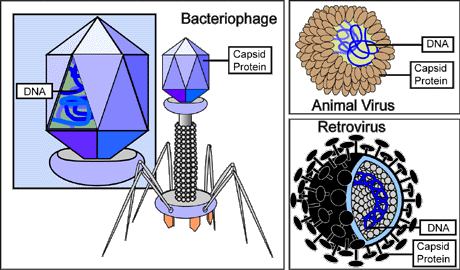
Figure 5. Types of Viruses
--------------------------------------------------------------------------------
This illustration depicts three types of viruses: a bacterial virus,
otherwise called a bacteriophage (left center); an animal virus (top right);
and a retrovirus (bottom right). Viruses depend on the host cell that
they infect to reproduce. When found outside of a host cell, viruses,
in their simplest forms, consist only of genomic nucleic acid--either
DNA or RNA (depicted as blue) surrounded by a protein coat, or capsid.
--------------------------------------------------------------------------------
There are three different ways genetic information contained in a viral genome can be reproduced. The form of genetic material contained in the viral capsid--the protein coat that surrounds the nucleic acid--determines the exact replication process. Some viruses have DNA, which once inside the host cell is replicated by the host along with its own DNA. Then, there are two different replication processes for viruses containing RNA. In the first process, the viral RNA is directly copied using an enzyme called RNA Replicase. This enzyme then uses that RNA copy as a template to make hundreds of duplicates of the original RNA. A second group of RNA-containing viruses, called the retroviruses, use the enzyme reverse transcriptase to synthesize a complimentary strand of DNA so that the virus's genetic information is contained in a molecule of DNA rather than RNA. The viral DNA can then be further replicated using the host cell machinery.
When the virus has taken over the cell, it immediately directs the host to begin manufacturing the proteins necessary for virus reproduction. The host produces three kinds of proteins: early proteins--enzymes used in nucleic acid replication, late proteins--proteins used to construct the virus coat, and lytic proteins--enzymes used to break open the cell for viral exit. The final viral product is assembled spontaneously, that is, the parts are made separately by the host and are joined together by chance. This self-assembly is often aided by molecular chaperones, or proteins made by the host that help the capsid parts come together.
The new viruses then leave the cell either by exocytosis or by lysis. Envelope-bound animal viruses instruct the host's endoplasmic reticulum to make certain proteins, called glycoproteins, which then collect in clumps along the cell membrane. The virus is then discharged from the cell at these exit sites, referred to as exocytosis. On the other hand, bacteriophages must break open, or lyse the cell in order to exit. To do this, the phages have a gene that codes for an enzyme called lysozyme. This enzyme breaks down the cell wall, causing the cell to swell and burst. The new viruses are released into the environment, killing the host cell in the process.
Why Study of Viruses?
Viruses are important to the study of molecular and cellular biology because they provide simple systems that can be used to manipulate and investigate the functions of many cell types. We have just discussed how viral replication depends on the metabolism of the infected cell. Therefore, the study of viruses can provide fundamental information about aspects of cell biology and metabolism. The rapid growth and small genome size of bacteria make them excellent tools for experiments in biology. Bacterial viruses have also further simplified the study of bacterial genetics and have deepened our understanding of the basic mechanisms of molecular genetics. Because of the complexity of an animal cell genome, viruses have been even more important in studies of animal cells than in studies of bacteria. Numerous studies have demonstrated the utility of animal viruses as probes for investigating different activities of eukaryotic cells. Other examples in which animal viruses have provided important models for biological research of their host cells include studies of DNA replication, transcription, RNA processing, and protein transport.
Deriving New Cell Types
Look closely at the human body and it is clear that not all cells are alike. For example, cells that make up our skin are certainly different from cells that make up our inner organs. Yet, all the different cell types in our body are all derived, or arise, from a single, fertilized egg cell through differentiation. Differentiation is the process by which an unspecialized cell becomes specialized into one of the many cells that make up the body, such as a heart, liver or muscle cell. During differentiation, certain genes are turned on, or become activated, while other genes are switched off, or inactivated. This process is intricately regulated. As a result, a differentiated cell will develop specific structures and perform certain functions.
Mammalian Cell Types
Three basic categories of cells make up the mammalian body: germ cells, somatic cells, and stem cells. Each of the approximately 100,000,000,000,000 cells in an adult human has its own copy, or copies, of the genome. The only exception being certain cell types that lack nuclei in their fully differentiated state, such as red blood cells. The majority of these cells are diploid, or have two copies of each chromosome. These cells are called somatic cells. This category of cells includes most of the cells that make up our body, such as skin and muscle cells. Germ line cells are any line of cells that give rise to gametes--eggs and sperm--and are continuous through the generations. Stem cells, on the other hand, have the ability to divide for indefinite periods and to give rise to specialized cells. They are best described in the context of normal human development.
Human development begins when a sperm fertilizes an egg and creates a single cell that has the potential to form an entire organism. In the first hours after fertilization, this cell divides into identical cells. Approximately four days after fertilization, and after several cycles of cell division, these cells begin to specialize, forming a hollow sphere of cells, called a blastocyst. The blastocyst has an outer layer of cells and inside this hollow sphere there is a cluster of cells called the inner cell mass. The inner cell mass cells will go on to form virtually all of the tissues of the human body. Although the inner cell mass cells can form virtually every type of cell found in the human body, they cannot form an organism. Therefore, these cells are referred to as pluripotent, that is, they can give rise to many types of cells but not a whole organism. Pluripotent stem cells undergo further specialization into stem cells that are committed to give rise to cells that have a particular function. Examples include blood stem cells that give rise to red blood cells, white blood cells and platelets and skin stem cells that give rise to the various types of skin cells. These more specialized stem cells are called multipotent-capable of giving rise to several kinds of cells, tissues, or structures.
The Working Cell--DNA, RNA, and Protein Synthesis
DNA Replication
DNA replication, or the process of duplicating a cell's genome, is required every time a cell divides. Replication, like all cellular activities, requires specialized proteins for carrying out the job. In the first step of replication, a special protein, called a helicase, unwinds a portion of the parental DNA double helix. Next, a molecule of DNA polymerase--a common name for two categories of enzymes that influence the synthesis of DNA-- binds to one strand of the DNA. DNA polymerase begins to move along the DNA strand in the 3' to 5' direction, using the single-stranded DNA as a template. This newly synthesized strand is called the leading strand and is necessary for forming new nucleotides and reforming a double helix. Because DNA synthesis can only occur in the 5' to 3' direction, a second DNA polymerase molecule is used to bind to the other template strand as the double helix opens. This molecule synthesizes discontinuous segments of polynucleotides, called Okazaki fragments. Another enzyme, called DNA ligase, is responsible for stitching these fragments together into what is called the lagging strand.
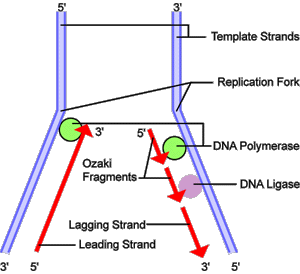
Figure 7. An Overview of DNA Replication
--------------------------------------------------------------------------------
Before a cell can divide, it must first duplicate its DNA. This figure
provides an overview of the DNA replication process. In the first step,
a portion of the double helix (blue) is unwound by a helicase. Next, a
molecule of DNA polymerase (green) binds to one strand of the DNA. It
moves along the strand, using it as a template for assembling a leading
strand (red) of nucleotides and reforming a double helix. Because DNA
synthesis can only occur 5' to 3', a second DNA polymerase molecule (also
green) is used to bind to the other template strand as the double helix
opens. This molecule must synthesize discontinuous segments of polynucleotides
(called Okazaki fragments). Another enzyme, DNA ligase (yellow) then stitches
these together into the lagging strand.
--------------------------------------------------------------------------------
The average human chromosome contains an enormous number of nucleotide pairs that are copied at about 50 base pairs per second. Yet the entire replication process only takes about an hour. This is because there are many replication origins sites on a eukaryotic chromosome. Therefore, replication can begin at some origins earlier than at others. As replication nears completion, "bubbles" of newly replicated DNA meet and fuse, forming two new molecules.
With multiple replication origin sites, one might ask how does the cell know which DNA has already been already replicated and which still awaits replication? To date, two replication control mechanisms have been identified: one positive and one negative. In order for DNA to be replicated, each origin of replication site must be bound by a set of proteins called the Origin Recognition Complex. These remain attached to the DNA throughout the replication process. Specific accessory proteins--called licensing factors--must also be present for initiation of replication. Destruction of these proteins after initiation of replication prevents further replication cycles from occurring. This is because licensing factors are only produced when the nuclear membrane of a cell breaks down during mitosis.
DNA Transcription--Making mRNA
DNA transcription refers to the synthesis of RNA from a DNA template. This process is very similar to DNA replication. Of course, there are different proteins that direct transcription. The most important enzyme is RNA polymerase--an enzyme that influences the synthesis of RNA from a DNA template. In order for transcription to be initiatied, RNA polymerase must be able to recognize the beginning sequence of a gene so that it knows where to start synthesizing an mRNA. It is directed to this initiation site by the ability of one of its subunits to recognize a specific DNA sequence found at the beginning of a gene--called the promoter sequence. The promoter sequence is a unidirectional sequence found on one strand of the DNA that instructs the RNA polymerase in both where to start synthesis and in which direction synthesis should continue. The RNA polymerase then unwinds the double helix at that point and begins synthesis of an RNA strand complementary to one of the strands of DNA. This strand is called the antisense or template strand, while the other strand is referred to as the sense or coding strand. Synthesis can then proceed in a unidirectional.
Although much is known about transcript processing, the signals and events that instruct RNA polymerase to stop transcribing and drop off the DNA template remain unclear. Experiments over the years have indicated that processed eukaryotic messages contain a poly(A) addition signal (AAUAAA) at their 3' end, followed by a string of adenines. This poly(A) addition, also called the poly(A) site, contributes not only to the addition of the poly(A) tail, but also to transcription termination and the release of RNA polymerase from the DNA template. Yet, transcription does not stop here. Rather, it continues for another 200 to 2000 bases beyond this site before it is aborted. It is either before or during this termination process that the nascent transcript is cleaved, or cut, at the poly(A) site, leading to the creation of two RNA molecules. The upstream portion of the newly formed, or nascent, RNA then undergoes further modifications, called post-transcriptional modification, and becomes messenger RNA (mRNA). The downstream RNA becomes unstable and is rapidly degraded.
While the importance of the poly(A) addition signal has been established, the contribution of sequences further downstream remains uncertain. A recent study suggests that a defined region--called the termination region--is required for proper transcription termination. This study also illustrated that transcription termination takes place in two distinct steps. In the first step, the nascent RNA is cleaved at specific subsections of the termination region, possibly leading to its release from RNA polymerase. In a subsequent step, RNA polymerase disengages from the DNA. Hence RNA polymerase continues to transcribe the DNA, at least for a short distance.
Protein Translation-- How Do Messenger RNAs Direct Protein Synthesis?
The cellular machinery responsible for synthesizing proteins is the ribosome. The ribosome consists of structural RNA and about 80 different proteins. In its inactive state, it exists as two subunits: a large subunit and a small subunit. When the small subunit encounters an mRNA, the process of translating an mRNA to a protein begins. In the large subunit, there are two sites for amino acids to bind, and thus be close enough to each other to form a bond. The "A site" accepts a new transfer RNA, or tRNA--the adaptor molecule that acts as a translator between mRNA and protein--bearing an amino acid. The "P site" binds the tRNA that becomes attached to the growing chain.
As we just discussed, the adaptor molecule that acts as a translator between mRNA and protein is a specific RNA molecule, the tRNA. Each tRNA has a specific acceptor site that binds a particular triplet of nucleotides--called a codon--and an anticodon site that binds a sequence of three unpaired nucleotides--the anticodon--which can then bind to the the codon. Each tRNA also has a specific charger protein; called an aminoacyl tRNA synthetase. This protein can only bind to that particular tRNA and attach the correct amino acid to the acceptor site.
The start signal for translation is the codon ATG, which codes for methionine. Not every protein necessarily starts with methionine, however. Oftentimes this first amino acid will be removed in later processing of the protein. A tRNA charged with methionine binds to the translation start signal. The large subunit binds to the mRNA and the small subunit, and so begins elongation--the formation of the polypeptide chain. After the first charged tRNA appears in the A site, the ribosome shifts so that the tRNA is now in the P site. New charged tRNAs, corresponding the codons of the mRNA, enter the A site and a bond is formed between the two amino acids. The first tRNA is now released and the ribosome shifts again so that a tRNA carrying two amino acids is now in the P site. A new charged tRNA then binds to the A site. This process of elongation continues until the ribosome reaches what is called a stop codon--a triplet of nucleotides that signal the termination of translation. When the ribosome reaches a stop codon, no aminoacyl tRNA binds to the empty A site. This is the ribosomes signal to break apart into its large and small subunits, releasing the new protein and the mRNA. Yet, this isn't always the end of the story. A protein will often undergo further modification, called post-translational modification. For example, it might be cleaved by a protein-cutting enzyme, called a protease, at a specific place or have a few of its amino acids altered.
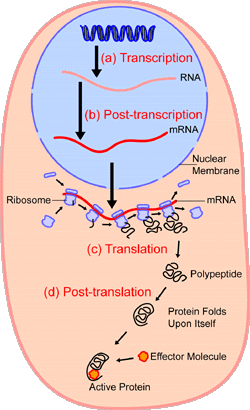
Figure 8. An Overview of Transcription and Translation
--------------------------------------------------------------------------------
This cartoon provides a graphic overview of the many steps involved
in transcription and translation. Within the nucleus of the cell (light
blue) genes (DNA, dark blue) are transcribed into RNA. This RNA molecule
is then subject to post-transcriptional modification and control, resulting
in a mature mRNA molecule (red) that is then transported out of the nucleus
and into the cytoplasm (peach) where it undergoes translation into a protein.
mRNA molecules are translated by ribosomes (purple) that match the three-base
codons of the mRNA molecule to the three-base anti-codons of the appropriate
tRNA molecules. These newly synthesized proteins (black) are often further
modified, such as by binding to an effector molecule (orange), in order
to become fully active.
--------------------------------------------------------------------------------
DNA Repair Mechanisms
Maintenance of the accuracy of the DNA genetic code is critical for both the long- and short-term survival of cells and species. Sometimes, normal cellular activities such as duplicating DNA and making new gametes, introduce changes or mutations in our DNA. Other changes are caused by exposure of DNA to chemicals, radiation, or other adverse environmental conditions. No matter the source, genetic mutations have the potential for both positive and negative effects on an individual as well as its species. A positive change results in a slightly different version of a gene that might eventually prove beneficial in the face of a new disease or changing environmental conditions. Such beneficial changes are the cornerstone of evolution. Other mutations are considered deleterious, or result in damage to a cell or an individual. For example, errors within a particular DNA sequence may end up either preventing a vital protein from being made or encoding a defective protein. It is often these types of errors that lead to various disease states.
The potential for DNA damage is counteracted by a vigorous surveillance and repair system. Within this system, there are a number of enzymes capable of repairing damage to DNA. Some of these enzymes are specific for a particular type of damage while others can handle a range of mutation types. These systems also differ in the degree to which they are able to restore the normal, or wild-type, sequence.
From Cells to Genomes
Understanding what makes up a cell and how that cell works is fundamental to all the biological sciences. Appreciating the similarities and differences between cell types is particularly important to the fields of cell and molecular biology. These fundamental similarities and differences provide a unifying theme, allowing the principles learned from studying one cell type to be extrapolated and generalized to other cell types.
Perhaps the most fundamental property of all living things is their ability to reproduce. All cells arise from pre-existing cells. That is, their genetic material must be replicated and passed from parent cell to progeny. Likewise, all multicellular organisms inherit their genetic information specifying structure and function from their parents. The next section of the genetics primer--What's In A Genome--details how genetic information is replicated and transmitted from cell to cell and organism to organism.
![]()