
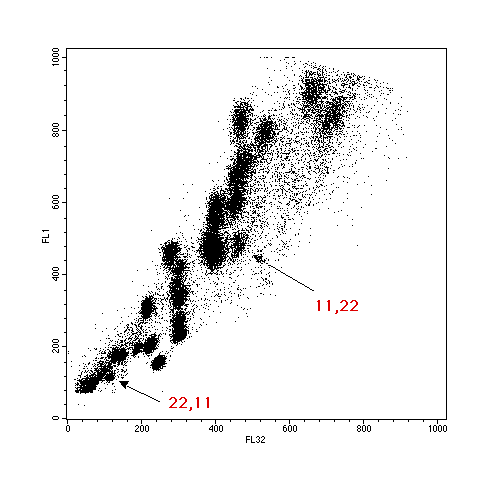
A good chromosome preparation is derived from a semi-confluent cell culture which is an exponential phase of growth. Cells are blocked in mitosis by the addition of colcemid. After an appropriate time (4 hours - overnight) the cells are disrupted using a detergent solution and the free chromosomes are recovered. The chromosome preparation is then stained with two DNA dyes - chromomycin A3 which stains CG-rich regions of DNA and Hoechst 33258 which stains AT-rich regions. A more detailed protocol can be found here. The chromosomes then separate according to their DNA content (size) and base pair composition. A bivariate plot shows clear separation of chromosomes 1-8, X, Y and 13-22, with only the group 9-12 not being individually defined as these are similiar in size and base pair composition.
Aberrations from normal profiles can be detected if there are sufficient differences - for example a trisomy 21 would be detected as a 50% increase in the chromosome 21 peak, or, in the case of translocations, chromosome size and base pair compostion may change sufficiently for a derivative chromosome to be clearly identified. This is an example of an 11,22 balanced translocation, where the two derivative chromosomes can clearly be identified.
Flow karyotyping has had only limited clinical use due to the polymorphism that occurs across the normal population.
The real advantage of flow cytometry is the utilisation of the sorting power of modern flow cytometers. Chromosome translocations and rearrangements that give an obvious population are able to be sorted and these sorted chromosomes are widely sought after within the human genetics field. Applications in this Institute include the preparation of genomic libraries, the preparation of chromosome paints from small numbers of sorted chromosomes and the mapping of genetic markers . This last application has been used, for example, to enable mapping of the break-off point in the 11,22 translocation, above.
{kind=link}
{kind=link}